



Споры о митохондриальной «Еве»
Лукавии же человецы и чародее преуспеют на горшее, прельщающе и прельщаемы. Ты же пребывай в нихже научен ecu и яже вверена суть тебе, ведый, от кого научился ecu
(2 Тим. 3, 13-14).
В данном докладе проводится общий анализ публикаций последних 18-ти лет в англоязычной научной литературе, в той или иной степени посвященных антропологическому феномену т.н. митохондриальной «Евы». Автор данного доклада, орнитолог по специальности, ставил своей целью рассмотреть данные, представленные по этому вопросу в наиболее значительных (ключевых) статьях и, не вдаваясь в технические детали, проанализировать их, как с точки зрения простой научной логики, так и православного представления об истории человечества.
В течение последних 20-ти лет, одной из самых динамично развивающихся отраслей биологии стала молекулярно-генетическая экология. Особенно это относиться к её разделу, занимающемуся выявлением взаимосвязей между молекулярно-генетическими особенностями популяций и происходившими в них реальными или гипотетическими демографическими и микроэволюционными процессами. Отношения между популяциями, основанные на их молекулярно-генетических сходствах и различиях, получили название филогенетических. Изучение межпопуляционных различий свидетельствует о том, что определённая генетическая дифференциация может быть обусловлена совокупностью экологических (например, географическая сегрегация) и демографическо-микроэволюционных факторов (например, историческая миграция, накопление точечных мутаций, эффект «бутылочного горлышка», «дрейф генов»), и могла существовать вполне реальная исходная родительская популяция. Однако, филогенетический подход стал активно применятся и к вопросам происхождения видов, т.е. вопросам, «макроэволюционным», несмотря на то, что никто никогда так и не объяснил каким именно образом микроэволюционные процессы могли бы привести к «макроэволюционным» изменениям и не нашёл предковых (промежуточных) видов существующих ныне форм [1, 2]. В частности, в последние годы молекулярно-генетические методы, в дополнение к традиционным морфометрическим, активно используются антропологами-эволюционистами в потугах установить процессы становления отдельных этнических и расовых групп людей, и Homo sapiens как вида.
Одним из ключевых методов филогенетики стало сравнение последовательностей нуклеотидных оснований различных локусов митохондриальной ДНК (мДНК) [3], т.е. ДНК находящейся в митохондриях – органеллах, отвечающих за внутриклеточную энергетику. Широкое использование именно этой молекулы ДНК в филогенетике обусловлено с одной стороны консервативностью её состава (число и характер кодирующих и некодирующих участков на уровне отрядов и ниже постоянен), а с другой стороны – более высокой, чем в ядерной ДНК, скоростью возникновения нейтральных точечных мутаций. Последнее качество способствует накоплению различий между изолированными популяциями (дивергенции), уже после нескольких тысяч или даже сотен поколений. Главным же свойством мДНК, делающим её приемлемой для филогенетического анализа, является её наследование у позвоночных практически исключительно по материнской линии, без рекомбинации отцовской и материнской генетической информации, как это происходит с ядерной ДНК. Таким образом, каждый индивид в данной генеалогической линии несёт копию мДНК, идентичною мДНК его матери (по сути – клон). В отсутствии рекомбинации, различия между линиями мДНК как сказано выше, накапливаются в следствии точечных мутаций. Таким образом, зная процент дивергенции мДНК (d) и скорость мутагенеза (s), можно вычислить теоретический период разделения (?) двух линий мДНК, представляющих отдельные группы особей (популяции), как d/s. Скорость мутагенеза (s), в свою очередь, равна d/t.
В 1987 г. журнал Nature опубликовал работу группы американских антропологов [4], вызвавшую оживлённую дискуссию, утихнувшую только недавно [5]. Авторы (Р. Канн и др.), проанализировав мДНК 147 людей – представителей коренных этносов Европы, Азии, экваториальной Африки, Новой Гвинеи и Австралии, представили следующие результаты: (1) африканцы, как популяция, обладают наибольшим разнообразием мДНК (0.47%, т.е. в среднем мДНК двух человек из данной популяции отличается на 0.47%); за ними следуют азиаты, австралийские и ново-гвинейские аборигены (0.35% и 0.25%) и европейцы (0.23%); (2) компьютерный анализ взаимоотношений типов мДНК, представленный как филогенетическое древо (см. ниже), систематически выделял часть африканцев в отдельную группу (ветвь). Эти результаты были интерпретированы как указывающие на Африку, как на первоисточник всех современных линий мДНК, т.е. на африканскую популяцию как самую древнюю, в следствии чего и накопившую наибольшее разнообразие типов мДНК; (3) после коррекции внутрипопуляционных различий, общий уровень дивергенции между популяциями, однако, был довольно низким (0.01-0.05%). Последнее наблюдение противоречило более ранней гипотезе, основанной на интерпретации археологических данных, о возможном независимом возникновении морфологически современных людей в нескольких географических регионах и их последующим смешением. Логическим следствием аналитического метода, использованного в этой публикации было предположение, что все ныне существующие линии мДНК могли произойти от линии одной женщины, впоследствии прозванной митохондриальной «Евой». Следует сделать оговорку, что здесь речь идет именно об одной, а не единственной женщине. Последнее, в принципе, неустановимо, так как при отсутствии потомков женского пола в одном из поколений, данная линия мДНК исчезает из популяции словно её и не было (по аналогии с исчезновением фамилии при отсутствии потомков мужского пола), хотя ядерная ДНК, несущая подавляющее большинство (99.99%) генетической информации, безусловно, передаётся. Так же, используя косвенные методы оценки скорости мутагенеза мДНК, авторы заключили, что предполагаемый исход из Африки произошел 140,000-280,000 лет тому назад.
Как и всякая работа, пытающаяся пролить свет на происхождение человека, статья Р. Канн и др. привлекла пристальное внимание как эволюционистов, так и креационистов. Уже в том же выпуске журнала Nature Дж. Уэйнскоут [6] указал, что, основываясь только
на митохондриальных данных, сложно сказать какая из двух групп популяций, африканская или евроазиатская, является предковой. Однако, в течении 4-х лет работа Канн пребывала в зените славы, и на нее активно ссылались, как на доказательство африканского происхождения современных людей около 200,000 «лет» тому назад. Беда началась в 1992 г. Г. Ги [7], ссылаясь на работу Темплтона [8] и сильно негодуя по поводу библейских параллелей, вызванных работой Канн, и которыми пестрела популярная пресса тех дней, показал, что статистически методы, использованные Р. Канн с коллегами, даже после дополнительной работы, опубликованной ими в 1991 г. в журнале Science (253: 1503-1507), не позволяют сделать абсолютно никаких выводов о том, какая из популяций является предковой. К такому же выводу пришёл и К. Уилле, после независимого анализа первичных данных Р. Канн и соавторов [9]. Эта критика была основана на трудностях в составлении и интерпретации филогенетических древ (напр., на выборе «правильного» древа и сотен тысяч возможных).
Филогенетическое древо, по своей концептуальной сути, гомологично «древу жизни», когда-то представленному Ч.Дарвином, и является не чем иным, как эволюционной гипотезой о временных отношениях современных таксонов (популяций, видов, семейств, и т.д.) между собой и с гипотетическими «общими предками». Причём не делается никаких различий между реальными микроэволюционными общими предками (уровень популяций и т.н. «видов-сестёр» (sister species)) и гипотетическими макро эволюционными общими предками (уровень родов, семейств и выше). Чем более сходны (в данном случае генетические) признаки таксонов, включённых в анализ, тем ближе их терминальные ветви и, соответственно, короче «эволюционное время» их разделения. Очевидно, что наблюдаемое здесь сходство общего плана строения, будь-то ДНК, скелет, поведенческие признаки или что-то иное, подменяется общностью происхождения. Эту подмену и сам принцип построения филогенетических древ можно проиллюстрировать следующим образом. Если мы сделаем 20 измерений (скажем длины, ширины, объёма, массы, числа составных элементов, наличия внешнего источника энергии, химического состава, и т.д.) популяций утюгов, молотков, гаечных ключей и трёх типов будильников, то, учитывая, что внутрипопуляционные различия слабее межпопуляционных, их самое простое общее филогенетическое древо может выглядеть следующим образом (Рис 1).
Будильники, в данном рисунке составляют общую («молодую») ветвь с короткими ответвлениями; ключи и молотки, составляют отдельную ветвь от будильников, но с более глубоким разветвлением, так как они менее сходны между собой, чем будильники (т.е. независимо «эволюционировали» более продолжительное время). И, наконец, утюги представляют отдельную, изолированную (самую «древнюю» в данной выборке) ветвь, обычно называемую «внешней группой» (outgroup). Позиции а, б, в и г представляют собой «общих предков» соответствующих терминальных ветвей. Эти «общие предки» именно так, т.е. чисто гипотетически, и обозначаются в публикациях в данной области, так как согласно «вере» эволюционизма, с её круговой логикой, «данные по современным видам являются единственным наличным и доступным свидетельством о «последнем общем предке»« (Дейнард и Кидд по [14]). (Прямо как в классическом примере, когда породы датируются по ископаемым, а ископаемые по породам).
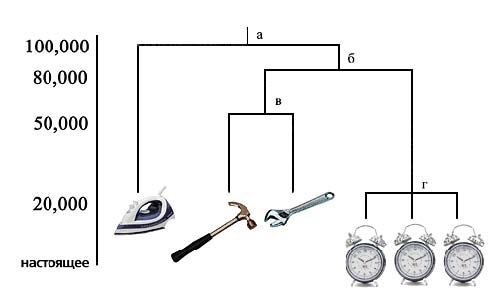
Рис.1. Филогенетическое древо предметов бытовой техники. Каждый символ соответствует отдельной популяции
Далее, если предположить, что разделение между линиями механической и электрической бытовой техники произошло, скажем, 100,000 «лет» назад (позиция «а»), то древо может быть снабжено хронологической шкалой, определяющей время появления отдельных ветвей и среднюю скорость накопления изменений. Обычно древо состоит из десятков и сотен образцов и, поэтому существуют миллионы возможных комбинаций ветвей и ответвлений. Чаще всего стремятся к составлению древ с самой простой структурой. В настоящее время также существуют методики определения статистической достоверности выбора структуры древа, основанного на конкретной выборке [3, 5].
Вернёмся к «Еве». Таким образом, на 1992 г. выводы Канн не были приняты и предполагалось, что, исторически, одинаковые типы мДНК могли существовать в разных популяциях даже после их географической сегрегации. А. Гиббонс [10], подводя черту под дискуссией на тот момент, заключила, что, хотя как гипотеза, африканская митохондриальная «Ева» ещё и не мертва, но по крайней мере смертельно ранена. Иными словами: данные просто слишком двусмысленны, чтобы свидетельствовать за или против африканского происхождения морфологически современных людей. На тот момент противники «африканской» гипотезы (напр. А. Темплтон) торжествовали. Тем не менее, в последствии, гипотеза «из Африки» была принята, основываясь, в том числе, и на работе К. Канн и др., и в настоящий момент является общепризнанной. Мы вернёмся, однако, к этому вопросу в конце доклада.
В дальнейшем дискуссия перешла с так и не разрешенного анализом мДНК вопроса «где жила «Ева»« к вопросу «когда она жила». Первоначальная оценка возраста «Евы» [4] была сделана косвенным образом на соотношении внутрипопуляционной дивергенции мДНК и предполагаемым возрастом колонизации континентов. Т.е., если заселение Нового Света произошло 12,000 «лет» назад и мДНК коренных американцев отличается в среднем на 0.04%, то долговременная средняя скорость мутагенеза в этой популяции =0.04/12,000. Из таких расчетов следовало, что мДНК мутирует со скоростью 2-4%/1 МЛ (миллион лет), что соответствует примерно одной мутации в 6000 поколений (одно поколение было принято за 20 лет). Учитывая, максимальный уровень дивергенции мДНК в выборке Р. Канн и др., предковая линия мДНК (собственно митохондриальная «Ева») должна была
существовать около 200,000 «лет» тому назад. Другие методики [11] также косвенным образом определяли скорость мутагенеза мДНК, основываясь на реальном отличии мДНК людей от шимпанзе (сравните, будильники и утюги, рис. 1) и гипотетическом периоде её накопления, начавшемся с появлением прямоходящих Homo и крупных приматов (сравните, позиция «а», рис. 1) в пластах, оцененных, согласно радиометрическим методам, в 5-7 МЛ. Эти калькуляции дали разброс в возрасте «Евы» от 70,000 до 800,000 «лет» [12]. Очевидно, что, помимо идеи существования общего предка «о» у человека и шимпанзе, эти оценки ещё основывались на идее постоянно тикающих «молекулярных часов». Этот униформистский принцип, аналогичный методам радиологической датировки пород, подразумевает, что мДНК мутирует с постоянной скоростью не только в разных популяциях и видах, но и в разных демографических и экологических условиях [3]. Поскольку выяснилось, что не только разные типы молекул ДНК, но и разные участки в молекулах одного типа мутируют с разной скоростью [3, 5], эта идея была подвергнута основательной критике (напр. [13, 19]). Очевидно, что для разрешения вопроса с датировкой «Евы», были необходимы прямые экспериментальные данные о скорости митохондриального мутагенеза. Такие данные были опубликованы группой Парсонса (здесь приводится по [12]) в 1997 г. Авторы, проведя анализ мДНК в 327 параллельных генеалогических парах (мать - ребёнок), обнаружили скорость мутагенеза, соответствующую 1 мутации кода контрольного участка мДНК в 30 поколений. Приблизительно такой же результат, 1 мутация в 40 поколений, был получен и группой Хауэлла (по [12]) годом раньше, но на основе более мелкой выборки (80 пар). Таким образом, реальная, непосредственно измеренная скорость мутагенеза человеческой мДНК была в 20 раз больше скорости, полученной косвенным образом. Из этих данных следует, что предковая линия всех современных типов мДНК существовала 6,000-6,500 лет тому назад, т.е. во времена Ноя и Великого Потопа.
Работа Парсонса и др., была принята с энтузиазмом учеными-креационистами. В частности, в 1998 г. К. Виланд [15] справедливо указал, что последние данные не доказывают исторического существования библейской Евы, поскольку наличие одной линии мДНК 6,000 лет назад не означает, что существовала всего одна женщина, но они ему и не противоречат. В тоже время они указывают на несостоятельность временных оценок эволюционистов.
Ответ не заставил себя долго ждать. А. МакЭндрю [16], отвечая непосредственно на статью Виланда, привёл ряд доводов против буквального принятия результатов Парсонса. Следует заметить, однако, что этот ответ напоминал диалог слепого с глухим, так как во главу угла МакЭндрю поставил утверждение, что возраст митохондриальной «Евы» в «6,500 лет определённо несовместим с известным возрастом современных людей» (выделено автором), т.е. с косвенными оценками этого возраста, основанными на эволюционистских принципах. В частности, ссылаясь на работу Хасегавы и др. [17], МакЭндрю указал, что некоторые мутации могут быть «слегка негативными», и потому присутствовать в генеалогических выборках, охватывающих всего один переход от поколения к поколению, но отсутствовать в популяционных, представляющих более долгий период «эволюции», в силу действия на них естественного отбора. Хотя принцип звучит логично, следует заметить, что работа Парсонса, как и Р. Канн и др., основана на анализе т.н. контрольного участка мДНК, в то время как Хасегава и др. анализировали кодирующие участки (сравниваются яблоки и апельсины). Анализ Хасегавы и др., и их выводы о «слегка негативных» мутациях, были основаны на косвенных данных. Более того, сами авторы признали, что экспериментально подтвердить их выводы «трудно». Конкретно, сравнивалось соотношение синонимичных (не влияющих на первичную структуру кодируемого белка) и несинонимичных (возможно негативно влияющих на структуру белка) мутаций. Поскольку внутри популяции это соотношение было в пользу
синонимичных мутаций, то было сделано предположение, что несинонимичные мутации слабее фиксируются в силу их отсеивания естественным отбором. Однако, во-первых, не было показано, что то, что верно для мутаций кодирующих участков, верно и для контрольного участка мДНК (поиск автором доклада публикаций на эту тему не принёс результатов). Во-вторых, разница в соотношении синонимичных и несинонимичных мутаций может просто означать различные скорости мутагенеза в разных участках мДНК [3]. Более того, вероятность фиксации даже «слегка негативных» мутаций в поколениях возрастает с уменьшением первоначального размера популяции [18], т.е. зависит от её демографической истории. Последний (2005 г.) обзор на тему мДНК и «эволюции» людей [19] указывает, что этот вопрос до сих пор окончательно не решён и, опять же, за отсутствием прямых данных.
Следующее возражение состояло в том, что контрольный участок мДНК не подходит для выявления «эволюционных» отношений популяций в силу различий в скорости мутагенеза в разных линиях, как было косвенно показано Ингманом и др., в 2000 г. [20]. Однако, совершенно определённо, что это возражение не имеет особого веса, так как ни учебник по молекулярной экологии [3], ни последний обзор по мДНК [19] ничего конкретного не говорят относительно неприемлемости контрольного участка для подобных исследований. Более того, именно контрольный участок мДНК наиболее активно используется в работах по филогенетике позвоночных и по сей день [3, 21]. Если же в действительности контрольный участок мутирует с разными скоростями в разных линиях мДНК только у людей, то вопрос возраста митохондриальной «Евы» в принципе на сегодня неразрешим без веры, либо в буквальное происхождение мДНК всех людей от Праматери Евы и «морфологически современных» людей от семьи Патриарха Ноя соответственно 7,500 и 6,000 лет тому назад, либо в происхождение линий людей и крупных приматов от загадочного «общего предка» 5-7 МЛ назад [17, 20, 22]. Прямые данные свидетельствуют скорее о первом.
Итак, единственные надежные выводы, которые можно сделать без натяжек и эволюционных догадок после 18-ти лет интенсивной и хорошо финансируемой работы, это то, что африканские народности обладают большим разнообразием мДНК, чем народы Азии, Европы и Нового Света [4, 20, 22] и что прямые измерения скорости мутагенеза мДНК не соответствуют гипотетической скорости их «эволюции». Также, по крайней мере некоторым исследователям (напр. [5, 14, 17]), стала очевидна круговая логика в попытках вывести демографическую историю современных людей из филогенетических данных, поскольку характеристики мДНК (следовательно и филогенетические древа) определяются именно этой историей, а не только промежутком времени в изоляции. В силу этого, Курно и Торн [14] заявили, что возможность сделать выводы об «эволюции» людей на основе анализа мДНК «ограничена», в то время как Пакендорф и Стоункинг [19] утверждают, что именно митохондриальные данные (т.е. работы Р. Канн, Ингмана, Мака-Мейер и др.) «подтвердили» африканское происхождение морфологически современных людей. Опять же, согласно [14] стартовая популяция современных людей была большой, в то время как большинство «экспертов» (напр., [23, 25]) полагают, что стартовая популяция была довольно маленькой – почти как в Деяниях, где Друзии оке убо и но нечто вопияху: бе бо собрание смущено, и множайшии от них не ведяху, чесо ради собрашася (Между тем одни кричали одно, а другие другое, ибо собрание было беспорядочное, и большая часть [собравшихся] не знали, зачем собрались. – А.М.) (Деян. 19,32). Примечательно так же то, что эволюционно-филогенетические древа людей (и обезьян), составленные на основе генетических и морфологических (обычно промеры частей скелета) признаков, не совпадают между собой [14, 25], указывая на разных «общих предков» и разные скорости «эволюции», и вынуждая эволюционистов прибегать
к новому нагромождению гипотез о «коэволюции» генетических, морфологических и поведенческих признаков [14,23,25].
В заключение хотелось бы сделать краткий экскурс в современные эволюционистские представления о происхождении человека и оценить их с точки зрения православного представления об истории человечества. На сегодняшний день мало у кого из антропологов-эволюционистов вызывает сомнение гипотеза, согласно которой первоначально большая часть Старого Света была заселена «Homo erectus», мигрировавшими примерно 2 МЛ назад из Африки. Относительно того, что произошло дальше, есть две версии. Согласно одной, основанной на интерпретации датировки и археологических данных, морфологически современные люди развились из «архаичных форм» независимо в разных регионах Старого Света и затем смешались. Этому противоречат генетические данные [4, 17, 20, 22]. Согласно другой, превалирующей в настоящее время гипотезе «недавнего африканского происхождения», морфологически современные люди вышли из Африки 100,000-200,000 «лет» тому назад и расселились в другие части света, вытеснив «Н. Erectus» и «H. Neanderthalensis», и не смешиваясь с ними [20, 23]. Проблемы с реконструкцией эволюционистами происхождения людей на основе морфометрических данных и радиометрической датировки окаменелостей достаточно хорошо известны (напр. [24]), признаются ими самими [14, 26] и не будут отдельно затрагиваться в данной работе. Здесь хотелось бы затронуть тему вытеснения «архаичных форм» и «недавнего африканского происхождения». Примечательны следующие наблюдения. Во-первых, генетическое различие между «неандертальцами» (прямой анализ последовательностей ядерной ДНК, Кавилли-Сфоза по [14]) и современными людьми такое же, как и между коренными жителями Новой Гвинеи и Таиланда. Во-вторых, согласно М. Вольпову (М. Wolpoff) (здесь приводится по [10]), морфологически, «неандертальцы» из Югославии похожи на европейцев, а «прямоходящие» из Китая и Индонезии на азиатов и австралийских аборигенов, соответственно. В третьих, период сосуществования «Н. Erectus» и «Н. Neanderthalensis» с современными людьми прекращается всего лишь около 20,000 «лет» тому назад [23], т.е во время максимального оледенения Евразии [26]. И, в-четвёртых, как указано выше, все построения о недавнем африканском исходе морфологически современных людей на основе генетических данных строятся на филогенетическом принципе: «чем сложнее, тем древнее», не имея никакой возможности принять во внимание демографическую историю народонаселения планеты (естественно, не обращаясь к Священному Писанию). Отбросив гипотетического общего макроэволюционного предка у людей и обезьян, безосновательную датировку эволюционистов и их разделение древних и современных людей на «виды», даже их собственные модели (напр. [5, 20, 22, 23]) согласны со следующим развитием событий. Около 6,000 лет назад население планеты состояло из стартовой популяции с небольшим числом линий мДНК, но большим разнообразием ядерной ДНК (т.е. семья Ноя, включавшая всего 3-х женщин – жён Сима, Хама и Иафета, имевших последующих потомков женского пола). Эта популяция существовала в районе Ближнего Востока/Передней Азии (т.е. место послепотопной высадки). Отсюда отдельные группы мигрировали в Африку (т.е. потомки Хама) и Евразию/Новый Свет (т.е. потомки Сима и Иафета). Африканская популяция, быстро размножившись в условиях тёплого, стабильного климата, в последствии не испытывала резких изменений численности [20] и, как следствие, сохранила разнообразие своей ДНК и даже в какой-то степени дифференцировалась за счёт накопления мутаций в мДНК и, видимо, ограниченного обмена генетической информацией с евро-азиатскими народностями (последнее может быть следствием проклятия Ноем потомства Хама). Послепотопное изменение климата привело к похолоданию и, возможно, оледенению в северных регионах Евразии, в результате чего целые этнические группы, по той или иной причине не мигрировавшие на юг, погибли или их численность резко снизилась. Такой феномен исторического
«бутылочного горлышка» широко распространён среди популяций животных и растений северного полушария, и связывается с оледенением их местообитаний [21, 27]. Эти процессы привели к уменьшению внутрипопуляционного разнообразия мДНК европейско-азиатских народностей и полному исчезновению отдельных линий («неандертальцы» и «прямоходящие») из современной популяции [5].
Очевидно, что доказать научно, насколько верна та или иная модель истории человечества в принципе невозможно, так как «правда всегда в глазах смотрящего», и интерпретация даже самых простых и очевидных фактов зависит сперва от веры и затем уже от знаний человека [24]. Хотелось бы только, чтобы мы чаще задумывались, что, веры всего две и, по слову Свят. Игнатия (Брянчанинова), только вера в правду и спасает и даёт возможность иметь здравое понятие о природе и человеке. Я завершаю этот доклад словами Свят. Григория Паламы [28], которые каждый Православный христианин, ученый или нет, может и даже должен привести себе на память, оказавшись в ситуации, когда данные, выдаваемые за научные, противоречат «здравому учению» Христовой Церкви: «Разве истина внешней науки не сомнительна и не смешана с ложью, почему всегда рано или поздно опровергается, что должны признать сами ее сторонники, тогда как другой истине по божественному слову Евангелия (Лк. 21, 15) никто не может противостоять, потому что она провозглашает очевиднейшую истину, ни в чем не смешанную с неправдой?... Так или иначе оказывается, что видов истины два: одна истина есть цель боговдохновенного учения, другую, не обязательную и не спасительную, ищет и никогда не находит внешняя философия».
Литература
1. Erwin, D.H. 2000. Macroevolution is more than repeated rounds of microevolution. Evolution and Development, 2:78-84.
2. Behe, M.J. 1998. Darwin's Black box: The Biochemical Challenge to Evolution. Touchstone, New York, NY.
3. Baker, A.J. 2003. Molecular methods in ecology. Blackwell Science Ltd. Oxford. UK.
4. Cann, R.L., Stoneking, M. & Wilson, A.C. 1987. Mitochondrial DNA and human evolution. Nature, 325:31-36.
5. Hagelberg, E. 2003. Recombination or mutation rate heterogeneity? Implications for mitochondrial Eve. Trends in Genetics, 19: 84-90.
6. Wainscoat, J. 1987. Out of the Garden of Eden. Nature. 325:13.
7. Gee, H. 1992. Statistical cloud over African Eden. Nature, 355: 583.
8. Templeton, A.R., Hedges, S.B., Kumar, S., Tamura, K., & Stoneking, M. 1992. Human origins and analysis of mitochondrial DNA sequences. Science. 255: 737-739.
9. Wills, C. 1992. Human origins. Nature, 356: 389.
10. Gibbons, A. 1992. Mitochondrial Eve: wounded, but not dead yet. Science, 257: 873-875.
11. Wills, C. 1995. When did Eve live? An evolutionary detective story. Evolution. 49:593-607.
12. Loewe, L. & Scherer, S. 1997. Mitochondrial Eve: the plot thickens. Trends in Ecology and Evolution, 12:422-423.
13. Howell, N., Elson, J.L., Turnbull, D.M. & Hernstadt, C. 2004. African haplogroup L mtDNA sequences show violations of clock-like evolution. Molecular Biology and Evolution, 21:1843-1854.
14. Curnoe, D. & Thorne, A. 2005. Number of ancestral human species: a molecular perspective. Homo. 53/3: 201-224.
15. Wieland, C. 1998. A shrinking date for 'Eve'. Сетевая версия.
16. MacAndrew, A. 2000. Misconceptions around mitochondrial Eve. Сетевая версия.
17. Hasegawa, M. Cao, Y. & Yang, Ziheng. 1998. Preponderance of slightly deleterious polymorphism in mitochondrial DNA: nonsynonymous/synonymous rate ratio is much higher within species than between species. Molecular Biology and Evolution, 15:1499-1505.
18. Woolfit, M. & Bromham, L. 2005. Population size and molecular evolution on islands. Proceedings of the Royal Society; B, 272: 2277-2282.
19. Pakendorf, B. & Stoneking, M. 2005. Mitochondrial DNA and human evolution. Annual Review of Genomics and Human Genetics, 6: 165-183.
20. Ingman, M., Kaessmann, H., Paa'bo, S. & Gyllensten, U. 2000. Mitochondrial genome variation and the origin of modern humans, Nature, 408: 708-713.
21. Buehler, D.M. & Baker, A.J. 2005. Population divergence times and historical demography in red knots and dunlins. Condor, 107:497-513.
22. Maca-Meyer, N., Gonzalez, A.M., Larruga, J.M., Flores, C. & Cabrera, V.M. 2001. Major genotnic mitochondrial lineages delineate early human expansions. BMC Genetics, 2:13.
23. Finlayson, C. 2005. Biogeography and evolution of the genus Homo. Trends in Ecology and Evolution, 20:457-463.
24. Rose, Fr. S. 2000. Genesis, Creation and early Man: The Orthodox Christian Vision. St. Herman of Alaska Brotherhood, Platina, CA.
25. Gura, T. 2000. Bones, molecules... or both? Nature. 406: 230-233.
26. Macdougall, D. 2004. Frozen Earth: The Once and Future Story of Ace Ages. University of California Press, Los Angeles, CA.
27. Hewitt, G.M. 1999. Post-glacial recolonization of European biota. Biological Journal of the Linnean Society, 68:67-112.
28. Христу, П. 2002. Двойственность познания по учению Святителя Григория Паламы. Сетевая версия.
